곤충은 대략 4억 8천만년전 공룡이 출현하기도 이전 시기인 고생대 석탄기 시기에 출현하였다. 그리고 21세기 현재까지 꾸준히 화석기록에서 모습을 나타내고 있으며, 현재는 모든 동물계 (animalia)에 속하는 생물 중에서 다양성이 가장 높다. 현재 곤충은 생태계에서 매우 다양하게 분포하며, 다양한 방식으로 생존하고 있다. 상당수의 곤충은 성장과정에서 번데기 과정을 거치는 완전변태 (Holometabolism)를 한다. 그외에 많은 종류의 개미 및 벌들은 꽃에서 꽃가루받이를 통해서 수분을 하거나, 사회군집을 만드는 진사회성 (Eusocial)으로 살아가기도 한다. 어떤 곤충은 다른 곤충의 번데기에서 기생(parasitoid)을 하며 살아가기도 한다. 다른 곤충을 흉내내는 의태 (Mimicry)나 주변과 비슷한 색깔로 위장(camouflage)을 하는 곤충도 존재한다. 곤충의 이런 다양성은 언제부터 시작된것인가에 대해서 화석기록을 통한 연구가 최근에 발표되었다.
1. 중생대 시기 곤충
중생대는 흔히 공룡의 시대라고 불리는 시대이다. 영화 쥐라기공원을 비롯한 많은 매체에서 공룡이 등장하는데 이 공료들이 실제로 살았던 시기가 중생대였기에 중생대를 공룡의 시대로 부르는 것이다. 그런데 중생대는 비단 공룡의 시대만은 아니었다. 곤충화석 기록을 보면, 곤충의 다양성이 폭팔적으로 증가하면서 동시에 현재까지 생태계에서 살아있는 곤충의 상당수가 중생대 시기에 나타났기 때문이다. 특히 완전변태를 하는 곤충, 진사회성 곤충, 꽃가루받이를 하는 곤충, 기생을 하는 곤충, 의태를 하는 곤충, 위장을 하는 곤충의 화석기록이 중생대 시기부터 모습을 드러내기 때문이다. 이증에는 분자배열 연구결과와도 일치하는 사례가 존재하기도 한다. 최근에 중국 난징의 과학 아카데미와 중국과학원 연구팀이 완전변태를 하는 곤충 (Holometabolous insect), 군집 생활을 하는 곤충 (Eusocial insect), 기생 생활을 하는 곤충 (Parasitoid insect), 꽃가루받이 생활을 하는 곤충(Polinating insect)에 대해서 발견되어온 화석기록을 리뷰한 논문을 발표하였다.
1. 곤충의 완전변태 (Holometabolous insect)
곤충 중에서 번데기과정을 보내는 곤충을 내시류 (Endopterygota)라고 한다. 이들은 곤충 전체 분류군 중에서 95%이상을 차지하는 매우 많은 분류군으로, 1) 환경 변화를 보다 더 잘 견딜수 있다는 점, 2) 유충으로 보내는 기간과 성충으로 보내는 기간을 나누어 상호간에 먹이 경쟁을 피할수 있게 하는 점을 근거로 곤충이 대규모로 번성할 수 있는 요인이 되었다.
번데기 과정을 곤충은 언제부터 시작한 것일까. 화석기록상 가장 오래된 기록은 석탄기 후기에 발견된 화석이다. 정확한 분류군은 알수 없으나, 번데기 과정을 거치는 곤충임은 분명하였다. 3억 1천 4백만 년 전 지층에서 발견된 곤충 아비오티에라 갈리카(Avioxyela gallica) -stem hymenopterid-, 스테파나스투스 폴리나에 (Stephanastus polinae) -stem coleopterid-, 그리고 메타볼라르바 벨라(Metabolarva bella)가 있다. 이중에서 아비오티에라와 스테파나스투스는 각각 오늘날 벌과 딱정벌레의 조상을 포함하는 Hymenopterid, Coleopterid에 속하는 것으로 보인다. 둘다 번데기 과정을 거치는 분류군이다.

하지만 완전변태를 하는 곤충의 본격적인 화석기록은 좀더 이후인 중생대 시기의 지층에서 나타난다. 프랑스의 북동부에 위치한 보주 산맥 (Vosges)에 위치한 2억 4천 7백만 년 전에서 2억 4천만 년 전 사이의 트라이아스기 지층인 볼치아의 사암 (Grès à Voltzia)에서 딱정벌레의 날개 화석이 발견되었으며, 중국 북동부의 트라이아스기 라디니알절인 2억 3천 8백만 ~ 2억 3천 7백만 년 전의 지층 (퉁촨층-Tongchuan Fm-)과 카르니안절 지층 (카라마이층-Karamay Fm-)에서 딱정벌레, 밑들이, 강도래, 물벌레의 화석이 발견되었다. 이중 딱정벌레, 밑들이, 강도래는 번데기 과정을 거치는 분류군이다.
한편 북미에서는 미국 유타주 남쪽의 트라이아스기 후기 카르니안절 시기에 만들어진 지층인 친리층 (Chinle Fm)에서 번데기 과정의 흔적화석이 구과식물 나무 (Conifer Wood)에서 발견되기도 하였다. 이 화석에는 실로크리프타 두로씨(Xylokrypta durossi)라는 학명이 붙었는데, 수직구조의 번데기방을 만든 점, 그리고 크기로 미루어보아 원시딱정벌레 (Archostemata)의 한 종류인 곰보벌레과(Cupedidae)의 곤충이 만든 것으로 추정된다고 한다. 비록 이 화석이 가장 오래된 내시류의 것은 아니겠지만, 이 화석은 가장 오래된 곤충의 번데기 흔적이 화석으로 남은 사례이다.
트라이아스기 시기는 후기에 접어들면서 기후의 극심한 변화가 일어났다. 대륙의 이동 및 해류의 흐름 변화등으로 인해 기후의 변화가 일어나면서 2백만년이 넘는 기간동안 대규모의 폭우가 쏟아지면서 트라이아스기 말 대멸종의 방아쇠를 당기게 되었다. 폭우가 끝나자 기후는 점차 건조해졌고 대멸종이 일어났는데, 번데기 과정을 거친 곤충은 이런 극심한 변화에서도 번데기 과정을 통해서 더 잘 버텨서 다른 곤충보다 더 많은 규모가 살아남았던 것으로 보인다.
2. 군집생활을 하는 곤충 (Eusocial insect)
인간은 사회 생활을 하는 사회적인 동물이라고 통칭한다. 그런데 이런 사회활동을 인간만 하는것이 아닌 곤충도 한다. 사회구조를 이루며 군집을 이루는 곤충을 사회성 곤충 (Social insects)이라고 하는데, 벌, 개미, 흰개미가 대표적으로 사회구조를 이루며 사는 곤충이다. 현재까지 발견된 군집생활을 하는 곤충 중에서 진딧물 (aphid), 총채벌레목 (thrips), 나무좀 (ambrosia beetles), 은주둥이 벌 (crabronid wasp), 어리호박벌아과(Xylocopine), 꽃등에아과(stenogastrine)의 경우에는 화석기록이 전무한 유령 분류군 (Ghost lineages)에 속한 상황이다. 이는 군집활동흔적이 퇴적활동에서 보존되기 어려우며, 이 곤충들중에서 진딧물, 총채벌레등은 지질학적으로 최근에 나타난 탓에 기록이 적은것으로 판단된다.
곤충의 군집활동에 대한 화석기록은 주로 흰개미, 개미, 벌에서 관측이 된다.
(1). 흰개미의 군집생활 화석기록
흰개미는 군집생활을 하는 곤충중 가장 오래된 곤충인 것으로 보인다. 화석기록과 분자배열 연구결과는 모두 흰개미의 기원이 대략 트라이아스기 후기 ~ 쥐라기 후기 시기에 나타난 것으로 추정된다. 다만 흰개미의 가장 오래된 화석기록은 호박에서 발견되는데, 미얀마에서 발견되는 1억년 된 호박에서 가장 오래된 흰개미의 화석이 여러 종 발견되었다. 2016년에 발견된 흰개미에는 크리쉬나테르메스 요드(Krishnatermes yoddh), 기간토테르메스 렉스(Gigantotermes rex)라는 학명이 붙었다. 기간토테르메스는 후에 기노르모테르메스속(Ginormotermes)으로 재분류 되었다. 2002년에 보고된 연구에서는 흰개미 스톨로테르메스과(Stolotermitidae)에 속하는 코스모테르메스 물투스(Cosmotermes multus)와 코스모테르메스 오파쿠스(Cosmotermes opacus)가 발견되었다. 코스모테르메스는 총 8개체의 병정개미와 56개체이 일개미, 25개체의 여러 모습의 어린 개체가 발견되었다. 화석이 발견된 퇴적환경을 미루어볼때 코스모테르메스는 눅눅하거나 썩은 나무 속에서 살았던 것으로 보이는데, 이는 이들의 군집생활 생태를 보여준다고 한다. 화석기록을 미루어보아, 흰개미는 최소한 1억년 전부터 군집을 이루며 여러 직위로 나누어진 생활을 하였던 것으로 보인다.
(2). 개미의 군집생활 화석기록
개미의 경우는 어떨까? 개미는 군집생활을 하는 곤충의 대표적인 예시이다. 이들은 한 둥지에서 어마어마한 생물량으로 군집을 이루며, 동시에 다른 곤충(ex: 진딧물)과 관계를 가지기도 한다. 이런 개미의 기원은 어떻게 되는가? 분자배열 연구 결과 개미는 쥐라기 후기에서 백악기 초기 시기에 벌의 한 부류에서 분화한 것으로 추정된다고 한다.
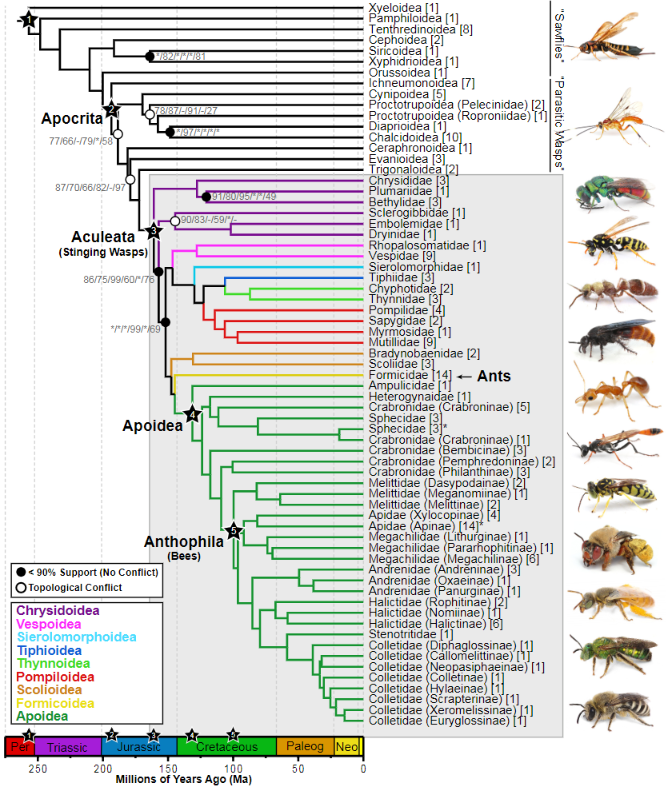
또한 화석기록에서도 가장 오래된 개미의 화석은 대략 1억년 전 즈음에 만들어진 미얀마의 호박에서 주로 발견되었다. 주로 지옥개미라고 불리는 하이도미르메스아과(Haidomyrmecinae)에 속하는 개미가 발견되었다. 지옥개미들은 특이한 점이 하나 있는데, 이들의 턱 구조는 오늘날 개미보다 척추동물과 더 비슷하게 움직인다는 것이다. 오늘날 개미들은 하악턱이 좌우로 움직이면서 먹이나 사물을 무는데, 과거 지옥개미들의 경우, 턱이 아래로 움직이는 방식을 하고 있다. 이런 턱구조는 개미보다는 척추동물과 비슷하다. 호박에서 주로 발견되는 지옥개미는 날개 없는 여왕, 날개 있는 수컷, 일꾼등으로 형태의 차이를 보이며 발견된다는 특징이 있다. 이는 개미들의 군집이 최소 1억년은 넘게 이루어졌음을 보여주는 근거라 할수 있다.

지옥 개미의 또 다른 특징은 다른 곤충이 개미에게 기생을 한다는 점이다. 오늘날에도 개미에 기생을 하는 곤충이 있는데, 예시중 하나로 딱정벌레중에서 풍뎅이붙이과(Histeridae)에 속하는 곤충이 있다. 이들은 개미를 꾀어내는 (myrmecophilous)곤충으로, 개미에 기생을 해서 침투한 뒤에 개미의 알이나 둥지 혹은 개미가 모아둔 먹이를 먹으며 산다. 이들은 주로 화학적, 혹은 형태적으로 개미를 속여서 기생을 한다. 이들은 주로 병정개미나 일개미들에게 기생을 하여서 개미의 둥지로 들어간다.

개미에게 기생을 하는 딱정벌레는 그 진화사가 1억년은 된듯 한다. 미얀마에서 발견된 호박에서 발견된 곤충 중에서 이렇게 개미에게 기생활동을 했던 것으로 추정되는 딱정벌레가 발견이 되었기 때문이다. 프로미르미스테르 키스트네리(Promyrmister kistneri)라는 곤충으로, 이 딱정벌레 역시 풍뎅이붙이과에 속한다. 이 곤충은 뒷다리 밑마디 (postcoxal)에 분비샘 (secretory glands)을 가지고 있는데, 이를 통해서 화학적 작용을 통해 개미의 둥지 내부로도 들어갈 수 있었을 것으로 추정이 된다. 개미는 보통 호르몬으로 의사소통을 하기에 개미에게 우호적인 호르몬을 분비하면 둥지로 들어가는것이 가능할 수 있기 때문이다. 이 딱정벌레는 개미의 군집생활이 최소 1억년은 되었음을 지시하는 또 다른 증거라 할 수 있다.
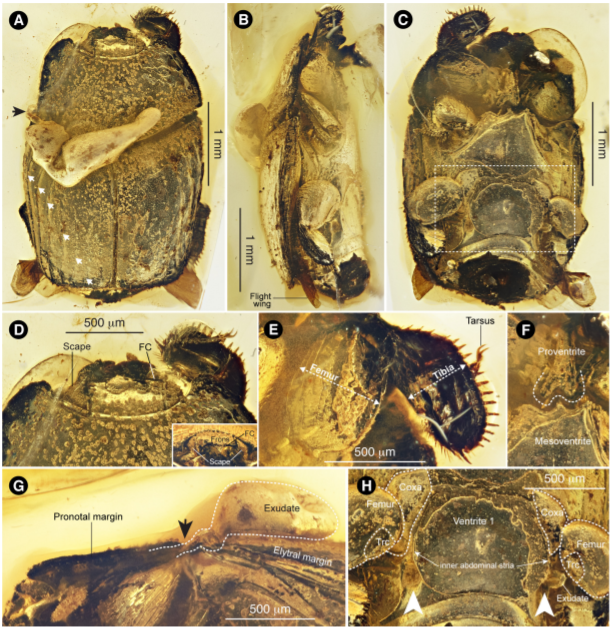
(3). 벌의 군집생활 화석기록
벌은 페름기 시기에 분화를 하였으며, 현재 2만여종이 살고 있으나, 군집생활은 꿀벌과 (Apidae), 꼬마꽃벌과(Halictidae)등등 따로따로 군집생활을 발달하였다. 그러나 다양성은 높지만 벌의 화석기록에서 군집생활에 대한 것은 개미나 흰개미와 달리 많지 않다. 벌의 군집생활이 남은 화석기록은 호박벌(Bumble bee), 꿀벌(Honey bee), 무침벌(Stingless bee)등에서 화석기록이 부분적으로 보일뿐이다. 대표적인 사례는 1988년에 보고된 미국 뉴저지주에 분포한 9천 6백만 년 전 즈음의 지층에서 발견된 호박에서 무침벌의 일종인 트리고나 피스카(Trigona pisca)의 사례가 있다.
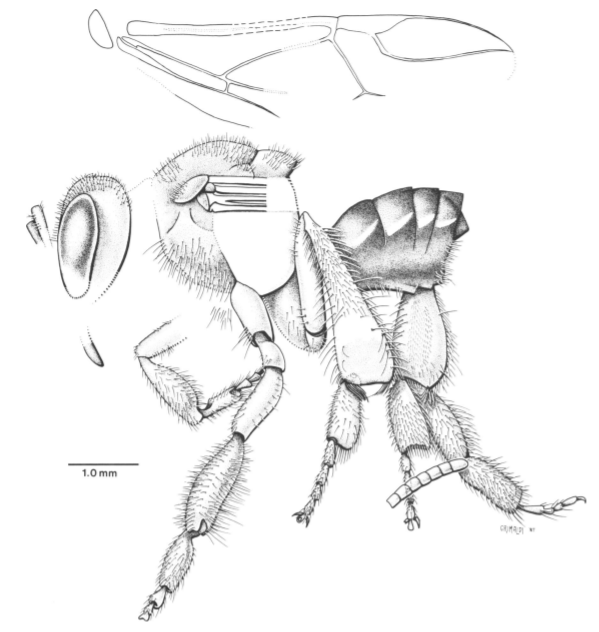
꿀벌이나 호박벌뿐 아니라 말벌, 그리고 쌍살벌과 호리병벌등이 속하는 말벌과(vespidae)의 경우 군집생활에 대한 화석기록이 희박하다. 중생대 시기 말벌의 둥지 화석이 미국 유타주에서 발견되었으나, 직접적으로 이들의 체화석은 중생대 이후인 팔레오세 때부터 발견이 되며, 그마저도 호리병벌은 화석기록이 부재한 상황이다. 종합해보면, 벌은 개미나 흰개미와는 달리 군집생활에 대한 화석기록이 부족한 상황이다.

3. 기생 생활을 하는 곤충 (Parasitoid insect)
오늘날 살아있는 곤충의 20%가 기생을 하며 살아간다. 현재까지 총 7개의 목-풀잡자리목(Neuroptera), 딱정벌레목(Coleoptera), 부채벌레목(Strepsiptera), 파리목(Diptera), 날도래목(Trichoptera), 나비목(Lepidoptera),벌목(Hymenoptera)-에서 기생활동이 관측된 바 있다. 이중에서 벌은 기생생활을 하는 곤충의 75%, 파리는 20%를 차지한다. 쥐라기 전기 이전의 곤충들의 기생생활에 대해선 뚜렷한 화석기록이 없어 알기 어려우나, 벌의 경우에는 쥐라기 전기서부터 화석기록이 등장한다. 따라서 최소한 쥐라기 전기서부터는 기생생활을 하였을 가능성이 있다고 받아들여진다.
기생생활을 하는 곤충은 주로 쥐라기 중기에서 백악기 전기동안 폭발적으로 증가한 것으로 보이는데, 당시 식물상의 변화로 영향을 받은 것으로 보인다.
4. 꽃가루받이 생활을 하는 곤충 (Polinating insect)
오늘날 꿀벌, 등에, 딱정벌레등 많은 곤충들이 꽃에서 꿀을 빨고 꽃가루를 옮기는 꽃가루받이 활동을 한다. 꽃의 정확한 기원 시점은 아직 알수 없지만, 화석기록을 보면 쥐라기 시기에 꽃이 살았던 것으로 보인다. 현재까지 발견된 가장 오래된 꽃은 중국 내몽골지역에서 살았던 플로리게르미니스 주라시카(Florigerminis jurassica)로, 이 꽃은 현재 존재하는 원시적인 꽃인 목련이나 암보텔라와 유사하다고 판단하였다. 꽃이 등장하기 전부터 살았던 식물인 겉씨식물의 경우에는 곤충의 꽃가루받이가 아니라 바람을 타고 꽃가루받이를 하였으리라고 판단된다.

그렇다면 곤충의 꽃가루받이는 언제부터였을까? 화석기록을 보면 꽃가루받이를 한 가장 오래된 곤충의 화석기록은 미얀마에서 발견된 호박화석에서 발견된 딱정벌레 안기모르델라 부르미티나(Angimordella burmitina)의 화석으로, 이 곤충의 화석은 꽃가루와 함께 발견되었다. 대략 9천 9백만 년 전에 살았던 이 곤충은 아직까지는 꽃가루받이를 한 가장 오래된 곤충의 사례로 뽑히고 있다. 딱정벌레 외에도 파리목, 밑들이목, 풀잠자리목, 총채벌레목(thysanoptera), 그리고 멸종한 분류군인 dictyoptera의 한 종류인 Alienopteridae에 속하는 곤충에서 꽃가루받이 흔적이 보인다고 한다.


중생대 시기 더듬이가 긴 곤충의 특이점이 하나 있는데, 겉씨식물에서 꽃가루 받이를 하는 곤충들은 다양성이 매우 높았다. 위에서 언급한 밑들이목과 풀잠자리목, 파리목은 백악기 중기 카친 호박에서 발견되는데, 총 5개의 과로 분류가 되며, 당시 백악기 중기에 꽃가루받이를 하는 곤충의 다양성과 복잡성이 높았다는 것을 보여준다. 당시 꽃의 꿀이 들어있는 관의 길이에 따라서 꽃가루받이를 하는 곤충들의 주둥이의 길이에 차이가 있었던 것으로 보인다.
5. 의태(mimicry), 위장(Camouflage)
의태와 위장은 생물이 포식자로부터 몸을 지킬때, 혹은 먹이를 사냥할 때 쓰는 수법이다. 이 패턴의 기원을 알기 위해서는 화석기록을 살펴보는 것이 필수적인데, 곤충의 일반적인 기원은 분자배열등으로 알수 있지만, 행동 패턴은 화석기록으로만 흔적이 나타나기 때문이다.
1. 의태
의태는 곤충이 다른 곤충이나 사물 (주로 식물)로 모습을 흉내내는 것으로, 그 정확한 기원이 고생대로 추정되지만 아직 그에 대해선 논란이 진행중이다. 의태에 대한 직접적인 화석기록은 중국 내몽골의 다오후고우(Daohugou)라거슈타테로, 그 시기는 대략 1억 6천 6백만 년에서 1억 5천 7백만 년 사이의 쥐라기 초기이다. 이 지역에서 풀잠자리류의 화석이 발견되었는데, 날개, 등의 형태가 식물과 유사하였다.
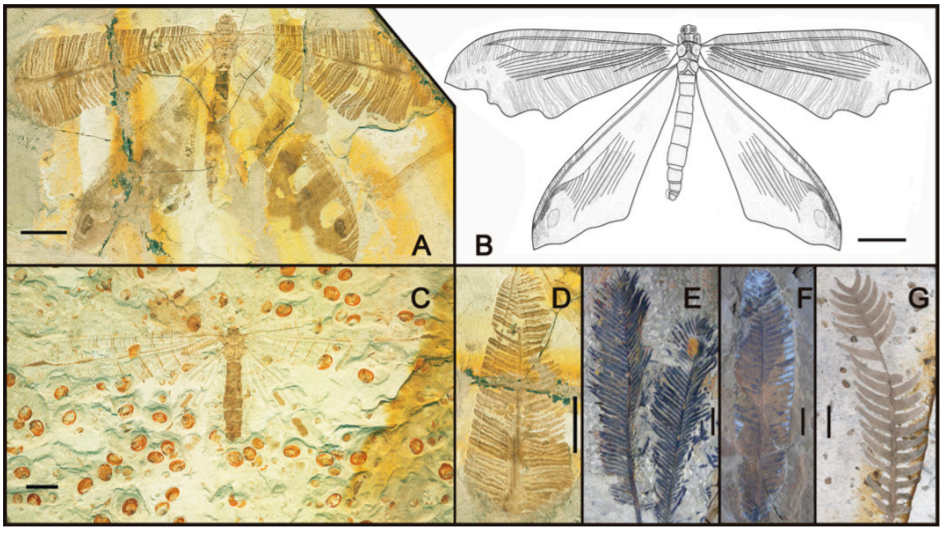
백악기 시기의 곤충화석에서도 이런 화석 기록이 등장한다. 브라질 북동부에 분포한 크라토 층 (Crato formtion)에서 잔가지를 닮은 형태의 대벌레를 닮은 메뚜기 프로스코피아과 (proscopiidae)에 속하는 에오프로스코피아 마르틸리 (Eoproscopia martilli)의 화석이 보고된 바 있다. 미얀마의 카친호박(Kachin amber)으로도 불리는 버마 호박(Burmese amber)에서는 좁쌀메뚜기과(Tridactylidae) 필론트리닥틸루스 왕기 (Phyllontridactylus wangi)의 화석이 발견된 바 있는데, 이 곤충은 우산이끼(liverwort)나 부처손과 식물인 셀라기넬라 (spike mosses)와 비슷한 형태의 중간다리 (mesothoracic legs)와 뒷다리 (metathoracic legs)를 가지고 있었다.

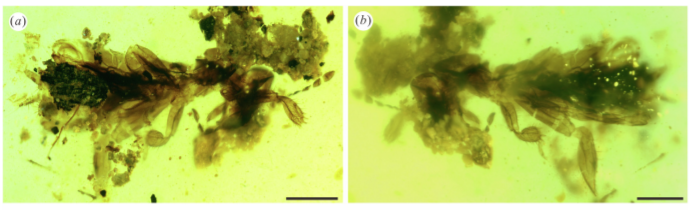
미얀마 호박에서는 좁쌀메뚜기뿐 아니라 풀잠자리의 유충과 대벌레의 화석이 보고된 사례도 존재한다. 풀잠자리상과(Chrysopoidea)에 속하는 필로크리사 후안기 (Phyllochrysa huangi)라는 풀잠자리의 유충, 그리고 대벌레목에서 정확한 과(family)를 알수 없는 대벌레 엑스타토소마 스티크툼 (Extatosoma stictum)가 발견된 사례가 있었다. 이 곤충은 가느다란 전흉배판(pronotum)과 다리 (leg), 복부 (abdomen)을 가지고 있었다.
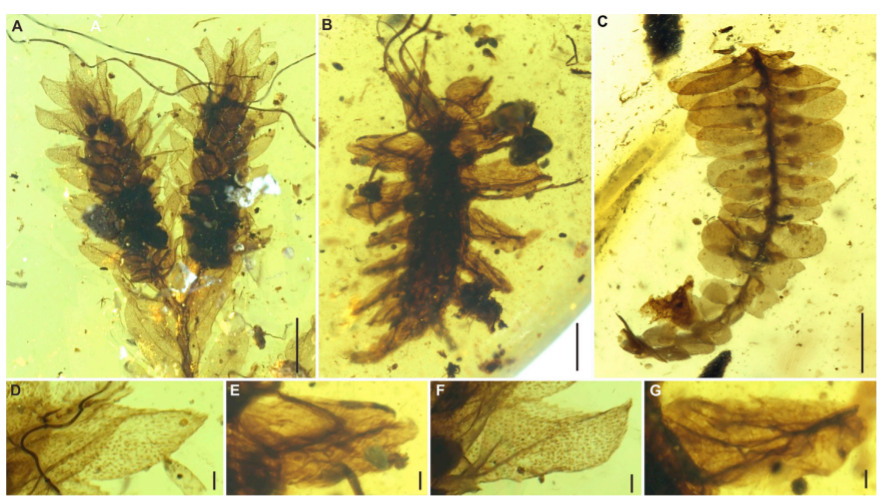

의태외에 곤충이 주변 환경에 맞게 위장을 하는 경우도 있는데 풀잠자리, 노린재는 주변환경에서 돌아다니는 물체를 이용해서 위장을 하는 행동이 관측되기도 한다. 화석기록에서 이런 사례는 많지는 않으나, 레바논, 스페인, 프랑스, 카친 호박에서 비슷한 행동 패턴이 관측된 사례가 있다.
카친 호박에서 발견된 곤충화석 중에서 다듬이벌레목(Psocodea)에 속하는 곤충과 두꺼비노린재과(Gelastocoridae)의 유충이 식물에서 떨어진 파편을 신체의 metanotum의 후방 경계면, 등, 복부의 abdominal tergum에 모아서 위장을 한 사례가 발견된 적 있다.

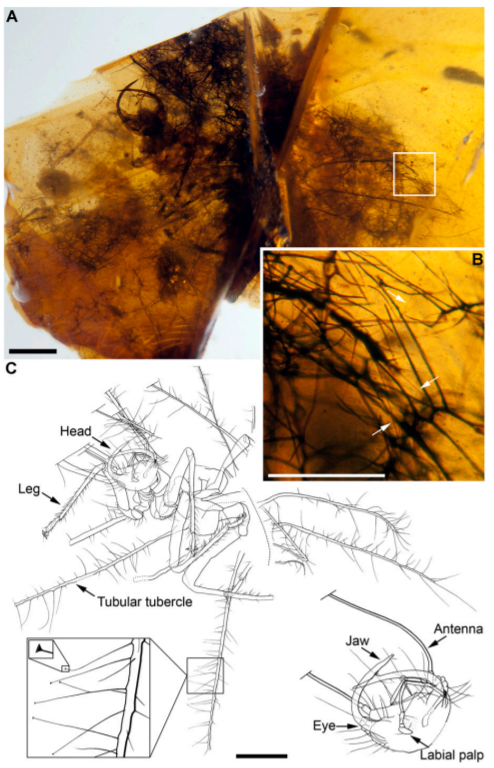
스페인에서도 곤충이 위장을 한 사례가 발견된 기록이 있다. 1억 1천 2백만 년 전에서 1억 9백만 년 전 사이 시기인 스페인 엘 소플라오 (El Soplao)지역에서 발굴되는 호박에서 뱀잠자리붙이상과 (Hemerobioidea)에 속하는 할루키노크리사 디오게네시 (Hallucinochrysa diogenesi)의 유충 화석이 보고되었다. 이 곤충의 유충은 등쪽에 몸을 덮을수 있는 관 형태이 결절 (tubular tubercle)을 아주 길게 가지고 있었다. 이 결절은 유충이 몸을 위장할 떄 필요한 부산물을 달아놓는 용도로 쓰이는 것으로 보인다. 이런 특징은 오늘날 풀잠자리 유충에서도 보이는 특징이다. 2018년에 레바논에서 발견된 1억 3천만 년 전에 쌓인 호박에서 티루스크리사 멜콰르트 (Tyruschrysa melqart)라는 풀잠자리의 유충 화석이 보고되기도 하였다. 이 유충은 흙을 몸에 붙여서 위장을 하였던 것으로 보인다.

의태와 위장은 곤충의 생존전략중 하나인만큼, 의태와 위장을 연구하는 것은 고환경 연구에 중요하다고 볼수 있다.
6. 곤충화석기록의 한계
곤충은 세계 여러 지역에서 서식하는 만큼, 화석도 세계 여러 지역에서 발견된다. 그중 절반 이상은 주요 화석지인 라거슈타테 (Lagerstätten)에서 발견되었다. 다만 화석으로 남기 어렵다는 작고 척추동물 대비 약한 신체 때문에 대부분의 곤충 화석은 분류학적으로 과(family)단계에서 다루어지는 경우가 대부분이다. 또한 호박처럼 곤충의 몸을 덮는 경우에는 소형 곤충의 화석도 보존되나, 진흙등으로 보존되는 퇴적환경에서는 소형 곤충의 호석이 보존되는 경우는 많지 않으며, 대부분 중형 이상의 곤충의 화석이 더 온전하게 보존되는 경우가 많다.
곤충의 화석은 보존되기 어려운만큼, 대멸종에서 어떻게 적응하여 살아남았는지 알기 어려운 경우가 많다. 특히 오늘날 곤충의 생태 대부분이 화석기록에서 보이는 중생대를 끝낸 K/Pg 대멸종에서 곤충이 어떻게 살아남아 진화하였는가는 알수있는것이 많지 않다.
곤충의 화석기록은 부족한 부분이 많은만큼, 현재까지 알려진 곤충의 진화사 상당수가 불완전한 부분이 많을수 있다는 것을 뜻한다.
연구 출처-
Wang, B., Xu, C., & Jarzembowski, E. A. (2022). Ecological radiations of insects in the Mesozoic. Trends in Ecology & Evolution.
사진 출처-
Bao, T., Wang, B., Li, J., & Dilcher, D. (2019). Pollination of Cretaceous flowers. Proceedings of the National Academy of Sciences, 116(49), 24707-24711.
Branstetter, M. G., Danforth, B. N., Pitts, J. P., Faircloth, B. C., Ward, P. S., Buffington, M. L., ... & Brady, S. G. (2017). Phylogenomic insights into the evolution of stinging wasps and the origins of ants and bees. Current Biology, 27(7), 1019-1025.
Chen, S., Yin, X., Lin, X., Shih, C., Zhang, R., Gao, T., & Ren, D. (2018). Stick insect in Burmese amber reveals an early evolution of lateral lamellae in the Mesozoic. Proceedings of the Royal society B: Biological Sciences, 285(1877), 20180425.
Cui, D. F., Hou, Y., Yin, P., & Wang, X. (2021). A Jurassic flower bud from the Jurassic of China. Geological Society, London, Special Publications, 521.
Engel, M. S., Barden, P., Riccio, M. L., & Grimaldi, D. A. (2016). Morphologically specialized termite castes and advanced sociality in the Early Cretaceous. Current Biology, 26(4), 522-530.
Heads, S. W. (2008). The first fossil Proscopiidae (Insecta, Orthoptera, Eumastacoidea) with comments on the historical biogeography and evolution of the family. Palaeontology, 51(2), 499-507.
Liu, X., Shi, G., Xia, F., Lu, X., Wang, B., & Engel, M. S. (2018). Liverwort mimesis in a Cretaceous lacewing larva. Current Biology, 28(9), 1475-1481.
Luo, C., Beutel, R. G., Engel, M. S., Liang, K., Li, L., Li, J., ... & Wang, B. (2021). Life history and evolution of the enigmatic Cretaceous-Eocene Alienopteridae: A critical review. Earth-Science Reviews, 103914.
Michener, C. D., & Grimaldi, D. A. (1988). The oldest fossil bee: Apoid history, evolutionary stasis, and antiquity of social behavior. Proceedings of the National Academy of Sciences, 85(17), 6424-6426.
Nel, A., Roques, P., Nel, P., Prokin, A. A., Bourgoin, T., Prokop, J., ... & Kirejtshuk, A. G. (2013). The earliest known holometabolous insects. Nature, 503(7475), 257-261.
Papier, F., Nel, A., Grauvogel-Stamm, L., & Gall, J. C. (2005). La diversité des Coleoptera (Insecta) du Trias dans le nord-est de la France. Geodiversitas, 27(2), 181-199.
Perrichot, V., Wang, B., & Barden, P. (2020). New remarkable hell ants (Formicidae: Haidomyrmecinae stat. nov.) from mid-Cretaceous amber of northern Myanmar. Cretaceous Research, 109, 104381.
Pérez-de la Fuente, R., Delclòs, X., Peñalver, E., Speranza, M., Wierzchos, J., Ascaso, C., & Engel, M. S. (2012). Early evolution and ecology of camouflage in insects. Proceedings of the National Academy of Sciences, 109(52), 21414-21419.
Pérez-de la Fuente, R., Peñalver, E., Azar, D., & Engel, M. S. (2018). A soil-carrying lacewing larva in Early Cretaceous Lebanese amber. Scientific Reports, 8(1), 1-12.
Tapanila, L., & Roberts, E. M. (2012). The earliest evidence of holometabolan insect pupation in conifer wood. PLoS One, 7(2), e31668.
Wang, Y., Liu, Z., Wang, X., Shih, C., Zhao, Y., Engel, M. S., & Ren, D. (2010). Ancient pinnate leaf mimesis among lacewings. Proceedings of the National Academy of Sciences, 107(37), 16212-16215.
Wenzel, J. W. (1990). A social wasp's nest from the Cretaceous period, Utah, USA, and its biographical significance. Psyche, 97(1-2), 21-29.
Xu, C., Wang, B., Fan, L., Jarzembowski, E. A., Fang, Y., Wang, H., ... & Engel, M. S. (2022). Widespread mimicry and camouflage among mid-Cretaceous insects. Gondwana Research, 101, 94-102.
Zhao, Z., Yin, X., Shih, C., Gao, T., & Ren, D. (2020). Termite colonies from mid-Cretaceous Myanmar demonstrate their early eusocial lifestyle in damp wood. National Science Review, 7(2), 381-390.
Zheng, D., Chang, S. C., Wang, H., Fang, Y., Wang, J., Feng, C., ... & Wang, B. (2018). Middle-Late Triassic insect radiation revealed by diverse fossils and isotopic ages from China. Science advances, 4(9), eaat1380.
Zhou, Y. L., Ślipiński, A., Ren, D., & Parker, J. (2019). A Mesozoic clown beetle myrmecophile (Coleoptera: Histeridae). Elife, 8, e44985.
'곤충 화석 연구' 카테고리의 다른 글
| 과 단위로 분석한 딱정벌레 화석기록에서 보이는 신종 출현과 멸종 빈도 (0) | 2021.08.26 |
|---|---|
| 진주층에서 발견된 두 방아벌레 화석 (0) | 2021.08.18 |




